恾侾丏傾僗僷儔僊儞懁嵔偺帺桼僄僱儖僊乕儅僢僾乮忋乯偍傛傃桳棙側椞堟偵偍偗傞戙昞揑側峔憿乮壓乯
DNA偲抈敀幙偲偺摿堎揑憡屳嶌梡
丂丂丂丂
恾侾丏傾僗僷儔僊儞懁嵔偺帺桼僄僱儖僊乕儅僢僾乮忋乯偍傛傃桳棙側椞堟偵偍偗傞戙昞揑側峔憿乮壓乯
丂堚揱巕偺敪尰偲偦偺惂屼偵偼丆偁傞墫婎攝楍傪摿堎揑偵擣幆偟寢崌偡傞抈敀幙偑廳梫側栶妱傪壥偨偟偰偄傞偲峫偊傜傟偰偄傞丅偟偐偟丆偦偺摿堎揑側擣幆偑偳偺傛偆側儊僇僯僘儉偱偍偙偭偰偄傞偺偐偵偮偄偰偼傢偐偭偰偄側偄丅偦偺婡峔傪柧傜偐偵偡傞栚揑偱丆扨偵憡屳嶌梡偺億僥儞僔儍儖僄僱儖僊乕傪媮傔傞偩偗偱偼側偔丆懁嵔偑條乆側峔憿傪偲傝偆傞偙偲傕峫椂偵偄傟偨帺桼僄僱儖僊乕儅僢僾偲偄偆奣擮傪採埬偟偨丅幚嵺偵儅僢僾傪寁嶼偡傞偙偲偵傛偭偰抈敀幙懁嵔偲妀巁墫婎懳偺摿堎揑側擣幆偼偙偺帺桼僄僱儖僊乕儅僢僾偐傜棟夝偱偒傞偙偲傪帵偟偨[1,2]丅
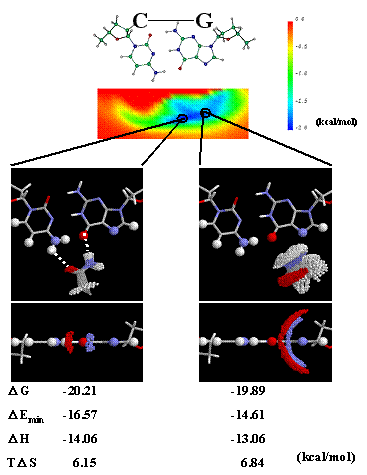
丂 墫婎懳偺major groove懁偱墫婎懳偺暯柺忋偵儊僢僔儏傪愗傝丆偦偺奿巕揰忋偵傾儈僲巁懁嵔偺Ca尨巕傪偍偔丅偦偺揰偵Ca尨巕偑偁傞応崌偵丆懁嵔偺峔憿傪宯摑揑偵敪惗偝偣傞偙偲偵傛偭偰偡傋偰偺峔憿傪栐梾偟丆偦傟傜偺僄僱儖僊乕傪寁嶼偡傞丅傾僗僷儔僊儞懁嵔偺応崌丆堦偮偺奿巕揰偱栺230枩揰偺峔憿傪敪惗偝偣傞丅奿巕揰偺悢偼栺500側偺偱丆堦偮偺儅僢僾偵偼栺10壄屄偺峔憿偺忣曬偑娷傑傟偰偄傞偙偲偵側傞丅恾侾偵偼丆C亅G墫婎懳偲傾僗僷儔僊儞懁嵔娫偺帺桼僄僱儖僊乕儅僢僾傪帵偡丅
丂 摑寁張棟傪巤偡偨傔偵偼旕忢偵懡悢偺峔憿傪敪惗偝偣丆偦傟傜偵懳偡傞僄僱儖僊乕寁嶼傪幚峴偡傞昁梫偑偁傞丅偟偐偟丆10壄屄偺僆乕僟乕偺ab initio MO朄寁嶼傪幚峴偡傞偙偲偼帠幚忋晄壜擻偱偁傞丅偦偙偱丆ab initio MO朄寁嶼傪嵞尰偡傞億僥儞僔儍儖僙僢僩傪偁傜偐偠傔嶌惉偟丆偦偺億僥儞僔儍儖娭悢傪巊偆偙偲偵傛偭偰丆ab initio MO朄寁嶼傪偟偨応崌偲摨偠寢壥偱偁傞偲婜懸偱偒傞寁嶼寢壥傪摼傞丆偲偄偆庤朄傪梡偄傞丅偄偔傜挿偄僔儈儏儗乕僔儑儞傪幚峴偟偰傕偦偙偱巊偭偨億僥儞僔儍儖偑怣棅偱偒側偄偺側傜偽偦偺寢壥偼怣梡偱偒側偄丆偲偄偆偙偲偼捈帇偡傋偒恀棟偱偁傞丅偙偙偱偺巹払偺戝検寁嶼偼丆妀巁偲抈敀幙娫偺憡屳嶌梡偺偨傔偵ab initio MO朄寁嶼偺寢壥傪僼傿僢僥傿儞僌偡傞偙偲偵傛偭偰嶌傜傟偨 ab initio potential [3] 傪巊梡偟偰幚峴偟偰偄傞丅
丂 恾侾乮忋乯偺儅僢僾偵偍偄偰丆惵偄椞堟偑丆傾僗僷儔僊儞懁嵔偲C亅G墫婎懳偲偺憡屳嶌梡偑帺桼僄僱儖僊乕揑偵桳棙側椞堟偱偁傞丅恾侾乮壓乯偵偼丆摨偠傛偆偵桳棙側椞堟忋偺2揰偵偮偄偰丆娭梌偡傞峔憿傪帵偟偨丅恾侾乮壓乯偵偍偄偰丆暘晍偟偰偄傞愒偄揰偼傾僗僷儔僊儞懁嵔偺僇儖儃僯儖婎偺巁慺尨巕丆惵偄揰偼傾儈僲婎偺拏慺尨巕丆敀偄揰偼悈慺尨巕偱偁傞丅恾侾乮壓乯偺嵍恾偲塃恾偲傪斾妑偡傞偲丆嵍懁偺曽偑億僥儞僔儍儖僄僱儖僊乕揑偵偼偐側傝埨掕側峔憿偱偁傞丅偟偐偟塃恾偺曽偑摨偠傛偆側埨掕惈傪帵偡峔憿偑暆峀偔暘晍偟偰偄傞丅偟偨偑偭偰帺桼僄僱儖僊乕揑偵偼摨偠傛偆側桳棙偝傪帵偟偰偄傞丅
丂偙偺傛偆偵丆帺桼僄僱儖僊乕儅僢僾傪昤偔偙偲偵傛傝丆扨偵嵟傕埨掕側峔憿傪摼傞偙偲偐傜摼傜傟傞傕偺偲偼堎側傞昤憸偑偁傜傢傟偰偔傞丅惗懱宯偺傛偆偵偝傑偞傑側梫場偑暋嶨偵偐傜傒偁偄側偑傜丆慡懱偲偟偰偁傞曽岦偵斀墳偑偡偡傫偱偄偔傛偆側宯傪棟夝偡傞偨傔偵偼丆偙偺傛偆偵乽摨偠傛偆側億僥儞僔儍儖僄僱儖僊乕傪傕偮懡偔偺応崌偑偁傝偆傞乿偲偄偆偙偲傪傆傑偊偨夝愅偑昁梫側偺偱偁傞丅
尨挊榑暥丗
[1] Fabio Pichierre, Misako Aida, M. Michael Gromiha and Akinori Sarai, "Free-energy maps of base-amino acid interactions for DNA-protein recognition," Journal of the American Chemical Society, 121, 6152-6157 (1999).
[2] Tomoki Yoshida, Tomohiro Nishimura, Misako Aida1, Fabio Pichierri, Michael M. Gromiha and Akinori Sarai, "Evaluation of free energy landscape for base-amino acid interactions using ab initio force field and extensive sampling," Biopolymers, 61, 84-95 (2002).
[3] Misako Aida, Giorgina Corongiu and Enrico Clementi, "Ab initio force field for simulations of proteins and nucleic acids," International Journal of Quantum Chemistry, 42, 1353-1381 (1992).
![]()